|
��
���Ľ� �ɿ��Ե��������ź�ת��
�ڶ���ķ����������漰���临�ӵ��ź�ת�����̣�����ǰ�漸���ᵽ���ź�;�����⣬���пɿ��Ե���ˮ����ص��ź�;������Wnt��Hedgehog��Notch��NF-��B���ź�;������Щ�ź�;������Ӱ������ϸ���ķֻ�����Ϊ�����źŷ��ţ�lateral signaling����
һ��Wnt�ź�;��
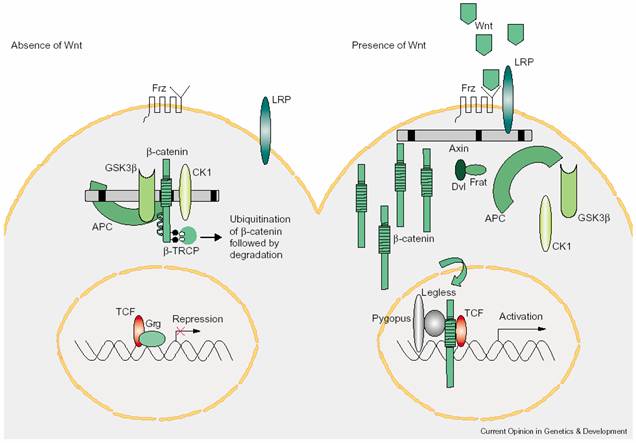
Wnt��һ��������ǵ��ף�ͨ���Է��ڻ��Է��ڷ������á���С���У���������������Wnt֮����������ٰ�������ΪInt1�������Ӭ�������Wingless��wg���и߶�ͬԴ�ԡ�Wnt�ź�;�������������-�������ף���-catenin�����ۡ���-catenin���ڹ�Ӭ�н�����������Armadillo����һ�ֶ�ܵĵ����ʣ���ϸ�����Ӵ������ճ������ã������γ�ճ�ϴ������������-catenin�ɽ���ϸ���ˣ����ڻ�����Wnt�ź��ڶ����������Ҫ���ã����쳣�����������������
Wnt�������Ǿ������ף�frizzled��Frz����Ϊ7�ο�Ĥ���ף��ṹ������G����ż�������壬Frz����N�˾��и������װ���Ľṹ��cysteine rich domain��CRD��������Wnt��ϡ�Frz�����������ڵ����ҵ��ף�Dishevelled��Dsh��Dvl����Dsh���ж���-catenin�Ľ���;�����Ӷ�ʹ��-catenin��ϸ�����л��ۣ�������ϸ��������Tϸ��������T cell factor / lymphoid enhancer
factor��TCF/LEF������������ڰл���ı��TCF/ LEF��һ�����˫����ڹ��ܵ�ת¼���ӣ�����Groucho������ƻ���ת¼����������-catenin��ٽ�����ת¼��Wnt����Ҫ����һ�����壨co-receptor������LRP5/6���������ܶ�֬����������ص��ף�LDL-receptor-related protein��LRP��������������������Frzһ��Dsh��
ͼ8-34 Wnt�ź�;�� ����Johan H. van ES 2003
Wnt�ź�;���ɸ���Ϊ��ͼ8-34����Wnt��Frz��Dsh����-catenin�Ľ��⸴�����ɢ����-catenin���ۣ�����ϸ������TCF/LEF������ת¼����c-myc��cyclinD1����
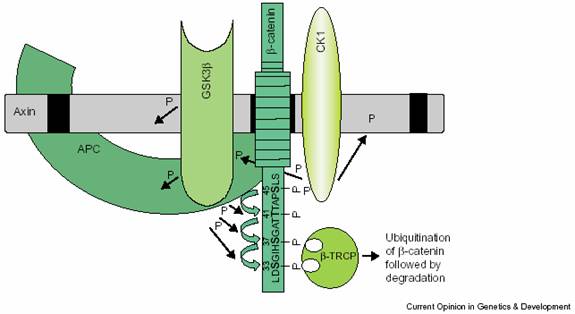
��-catenin�Ľ��⸴���壺��Ҫ��APC��Axin��GSK-3����CK1�ȹ��ɡ�
GSK-3������һ�ֵ���ø����û��Wnt�ź�ʱ��GSK-3���ܽ�������żӵ���-catenin�����˵�˿����/�հ���л��ϣ����ữ����-catenin�ٽ�ϵ���-TRCP���������ܷ����Ĺ������Σ�������ø�壨proteasome�����⡣��-catenin�б�GSK3���ữ�İ��������г�Ϊ�ƻ��У�destruction box���������з��������������ijЩ��֢��
CK1���ҵ���ø��casein kinase 1�����ܽ���-catenin��Ser45���ữ�����GSK-3������-catenin��Thr41��Ser37��Ser33���ữ��ͼ8-35����
ͼ8-35 ��-catenin�����ữ ����Johan
H. van ES 2003
APC����һ���ְ�������ͻ�������������������᳦������Ϣ����adenomatous polyposis coli����������ʱ������ƣ����ܷ�����䡣APC������������ǿ���⸴��������-catenin��������
Axin����һ��֧�ܵ��ף����ж���������������õ�λ�㣬�ܽ�APC��GSK-3������-catenin��CK1�����һ�𡣴�����������Dishevelled��PP2A��protein phosphatase 2A���ȳɷֽ�ϣ�����Dsh��Axin�����ʹ���⸴������塣PP2A��������Axinȥ���ữ����ʹ���⸴������壬�������Wnt;�������������ӣ���PP2A�����ɴ��ǻ��͵����ǻ������ֹ��ɣ�������ǻ����������ְ�����
Wnt�ź�;���������ɷ֣�
GBP��GSK-3����ϵ��ף�Frat����IJ������Wnt�ź�;�������������ã�GBP/Frat����GSK3-���Ļ��ԡ�
Dickkopf1��DKK1������һ�ַ��ڵ��ף�����Wnt����LRP5/6����һ�ഩĤ����Kremen1/2��ϣ��γ������壬�յ����ٵ�ϸ�����̣�����ϸ��Ĥ�ϵ�LRP5/6���ɴ������Wnt�ź�����ڵĴ��ݡ�
sFRP��������Frz��ص��ף�secreted frizzled-related
proteins��������һ��CRD�ṹ��ȱ���ߴο�Ĥ����������Frz�������Wnt���ס����������Ƶ�����Sizzled��WIF-1��Cerberus������Ҳֱ����Wnt����ϣ��Ӷ���Wnt�źš�
����Notch�ź�;��
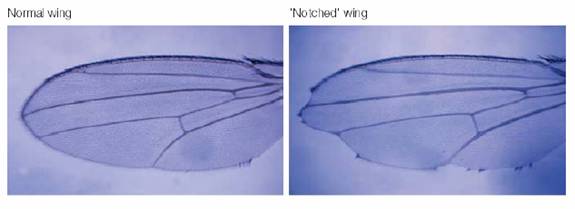
Notch�������緢���ڹ�Ӭ�����ֹ���ȱʧ���³�Եȱ�̣�notches��ͼ8-36��������̥�����У�����Ƥ��֯��ǰ��ϸ���зֻ�����Ԫϸ������ϸ������Notch����Delta������ϸ��Ĥ�ϵ�Notch��ϣ������ź�;������ֹ����ϸ������ͬ���ķֻ���������������������ƣ�lateral inhibition����Notchͻ��İ������������̥������������̥������֯ȡ������Ƥ��֯�Ӷ�ʹ����֯�쳣�ḻ��
ͼ8-36 Notchȱ�������Ӭ��Եȱ��
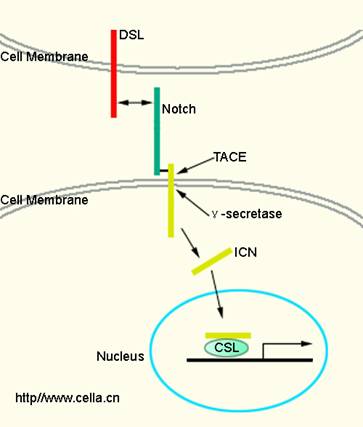
Notch�ź�;����Notch��Notch���壨DSL���ף���CSL��һ��DNA��ϵ��ף�����ɡ�Notch���������Ϊ���ο�Ĥ���ף������壨��Delta��������ϸ����Notch��Ϻ�Notch������ø���и�ͷų����к˶�λ�źŵİ�����ICN��intracellular domain of Notch��������ϸ������CLS��ϣ����ڻ������ɸ���Ϊ��ͼ8-37����Delta��Notch��ø����ICN������ϸ������CLS-ICN������������ת¼��
Notch��Ϊ������Լ300KD�ĵ����ʣ���Ӭֻ��1��Notch��������4����Notch1-4����Notch�İ������ǽ������������в�ͬ������EGF���ظ����У�EGF-R����3��LNR��Lin/Notch repeats������������RAM��RBP-J kappa associated molecular���ṹ��6��ê���ף�cdc10/ankyrin��ANK���ظ����С�2���˶�λ�ź���NLS����PEST�ṹ��RAM�ṹ������CSL��ϵ�����PEST�ṹ����Notch�Ľ����йء�Notch����Ҫ���������и��һ���ڸ߶������ڱ�furin�и�Ϊ2��Ƭ�ϣ�ת�˵�ϸ��Ĥ�γ�������塣�������ϵ���������Notch�����ַ������ζ��ѣ����DZ�������������-��-ת��ø��TNF-��-converting enzyme��TACE���иȻ����-�ٷ���ø����-secretase���и������Ҫ���ϵ��ף�presenilin��PS�����롣ø���Ժ��ͷ�Notch������ICN������ϸ���˷�������ѧ���á�
Notch���壺�ڹ�Ӭ��Notch������ΪDelta��Serrate���߳�ΪLag-2��ȡ��д��ĸ��Notch�������ֱ���ΪDSL���ף��ڲ��鶯���н���Jagged�������ǵ��ο�Ĥ�ǵ��ף�������������������ȵ�EGF���ظ�����N����һ�����Notch������DSL����
CSL��Ϊת¼���ӣ��ڲ��鶯���н���CBF1���ڹ�Ӭ�н���Suppressor of Hairless�����߳��н���Lag-1��������CSL��ʶ����ض���DNA���У�GTGGGAA�����������λ��Notch�յ�������������ϡ�ICN������ʱ��CSLΪת¼�������ӡ������ICNʱ��CSL���յ���ػ���ı��Notch�źŵİл����Ϊ��������-��-������ת¼���ӣ�basic helix-loop-helix��bHLH���������ֵ���������ϸ���ֻ�ֱ����صĻ����ת¼���粸�鶯���е�HES��hairy/ enhancer of split������Ӭ�е�E��spl�� ��enhancer of split��������צ��е�XHey-1�ȡ�
ͼ8-37 Notch�ź�;��
����Hedgehog�ź�;��
Hedgehog��һ�ֹ��۽�ϵ��̴��ķ����Ե��ף��ڶ����������Ҫ���á���Ӭ�ĸû���ͻ�䵼���׳�������������ͻ�����ƴ�⬣�����Hedgehog����������������3���������Hedgehog���ף�����Shh��Sonic hedgehog����Ihh��Indian hedgehog����Dhh��desert hedgehog��������Shh�Ǹ��ݵ�����Ϸ�еĽ�ɫ�����ģ����������ô�⬵������������ġ�
������Ĥ����Patched��Ptc����Smoothened��Smo���鵼Hedgehog�ź�����ڴ��ݡ�Ptc��12�ο�Ĥ���ף�����Hedgehog��ϣ�SmoΪ7�ο�Ĥ���ף���G����ż��������ͬԴ������Hedgehog������£�Ptc����Smo����Hedgehog��Ptc���ʱ��������Ptc��Smo���������ã����������¼���ͼ8-38����
Hedgehog�ź�;����ת¼������Ci��Cubitus interruptus���ڼ�������ΪGli��������пָ�ṹ��������155KD���ڰ�����Ci�����������γɸ����壬��Щ���װ�����Fu��Fused��һ��˿����/�հ��ἤø����Cos��Costal��һ���ܽ�������ê�������ϵĵ��ף���Su��suppressor of Fused�����䵰�ף�����û��Hedgehog�ź�ʱ��Ci��ˮ��Ϊ75KD��Ƭ�Σ�����ϸ���ˣ�����Hedgehog�ź���Ӧ����Hedgehog��Ptc���ʱ��Ci�Ľ��ⱻ���ƣ��Ӹ��������ͷų�����ȫ����Ci����ϸ�����У�������ػ�������Щ�������Wnt��Ptc��Ptc�ı���ֻ�����Smo���Ӷ�����Hedgehog�źţ���һ�ַ������ڡ�
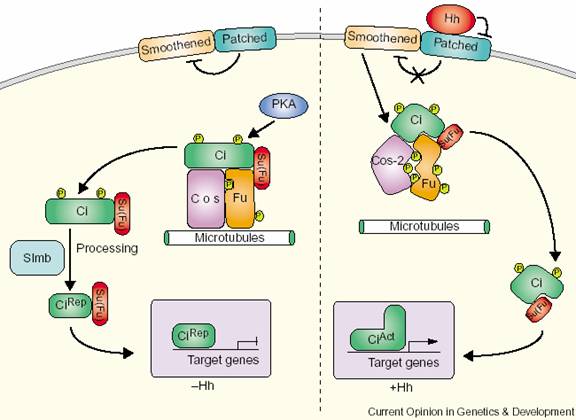
ͼ8-38 Hedgehog�ź�;�� ����Kent Nybakken��Norbert
Perrimon 2002
�ġ�NF-��B�ź�;��
NF-��B������Rel�����ת¼���ӣ����������������ߡ���֢��Ӧ��ϸ���ֻ��йصĻ���ת¼�����鶯��ϸ����������NF-��B/Rel��RelA(P65)��RelB��C-Rel��NF-��B1(P50)��NF-��B2(P52)��������RelͬԴ��(Rel homology domain��RHD)�����γ�ͬ��������壬������ͬ�Ļ���ת¼����Ϣ״̬�£�NF-��B�����������Ƶ���I��B��ϳ����������������ϸ���ʣ�����̼��ɼ���I��B�ķ��ػ�����;������ʹNF�C��B�����������ˣ����ڻ���ת¼��
I��B�����Ա��I��B����I��B����I��B����I��B����I��B����Bcl-3�ȣ���������Rel��������õ�ê�����ظ����к��뽵���йص�C��PEST���С�
IKK(I��B
kinases)��NF-��B�źŴ���ͨ·�Ĺؼ��Լ�ø�������ź��磺����������������tumor necrosis factor��TNF��������1(interleukin-1��IL-1)�ȿ��Լ���IKK��ʹI��B���ữ������ػ�;�����⡣
����ӣ�hemizygote������Ȼ�Ƕ����壬��һ�����������ǵ��۵ģ�û����֮���Ӧ�ĵ�λ������
|