|
ΓΓ
ΒΎ»ΐΫΎ
ΡΘ ΫΕ·ΈοΓΣΓΣΙϊ”§
œ÷‘ΎΈ“Ο«Ω…“‘±»Ϋœ«εΈζΒΡΩ¥ΒΫ ήΨΪ¬―ΖΔ”ΐΈΣ–¬ΗωΧεΘ§ « ή“ΜœΒΝ–Μυ“ρΒςΩΊΒΡΘ§’β–©Μυ“ρ‘ΎΖΔ”ΐΙΐ≥Χ÷–Θ§Α¥’’ ±ΦδΓΔΩ’ΦδΥ≥–ρΤτΕ·ΚΆΙΊ±’Θ§ΜΞœύ–≠ΒςΘ§Ε‘≈ΏΧΞœΗΑϊΒΡ…ζ≥ΛΚΆΖ÷Μ·Ϋχ––ΒςΫΎΓΘœΗΑϊΖ÷Μ·ΒΡ Β÷ «‘ΎΆβΫγ–≈Κ≈ΒΡΉς”Οœ¬Θ§ΑϊΡΎΚΥ÷ œύΜΞΉς”ΟΒΡΫαΙϊΓΘ
ΗυΨίΕ‘Ιϊ”§Β»ΡΘ Ϋ Β―ιΕ·ΈοΒΡ―–ΨΩΘ§ΖΔœ÷‘Ύ¬―œΗΑϊ÷–Θ§ Ήœ»±μ¥οΒΡ «ΡΗΧεΜυ“ρΘ§’β–©Μυ“ρ‘Ύ‘ΎΉΧ―χœΗΑϊ÷–ΉΣ¬ΦΘ§»ΜΚσ±Μ δ»κ¬―œΗΑϊΘ§»γbcdΓΔhbΓΔnosΓΔcdlΒ»Μυ“ρΓΘ’β–©Μυ“ρ≤ζΈο‘Ύ≈Ώ–Έ≥… ±Θ§―Ί«ΑΚσ÷α–Έ≥…“ΜΗω≈®Ε»ΧίΕ»Θ§ΨωΕ®ΝΥ≈ΏΒΡ«ΑΚσΈΜ÷ΟΚΆΆΖΈ≤«χ”ρΘ§’β–©ΡΗΧεΜυ“ρΒΡ≤ζΈο «“Μ÷÷DNAΫαΚœΒΑΑΉΦ¥ΉΣ¬Φ“ρΉ”Θ§ΥϋΦΛΜνΖ÷ΫΎΜυ“ρΒΡΉΣ¬ΦΘ®ΆΦ14-10Θ©ΓΘ
BicoidΘ®bcdΘ©ΚΆHunchbackΘ®hbΘ©ΨωΕ®≈ΏΧΞ«ΑΕΥΒΡ–ΈΧ§Θ§bcdΜυ“ρ ßΜνΜα“ΐΤπΆΖ–Ί≤Ω»± ßΘ§≤Δ ΙΆΖ«χ±δΈΣΈ≤ΫΎΒΡΧΊ’ςΓΘNanosΘ®nosΘ©ΚΆCaudalΘ®cdlΘ©Β»ΨωΕ®ΚσΕΥΒΡ–ΈΧ§Θ§TorsoΘ®torΘ©ΨωΕ®≈ΏΧΞΡ©ΕΥΒΡ–ΈΧ§Θ§’β–©Μυ“ρΙ≤Ά§ΨωΕ®≈ΏΧΞΒΡ«ΑΚσ÷αΓΘ
≈ΏΧΞΒΡ±≥ΗΙ÷α”…20ΕύΗωΜυ“ρΨωΕ®Θ§Τδ÷–DorsalΘ®dlΘ©ΆΜ±δΜα“ΐΤπ≈ΏΧΞ±≥≤ΩΜ·Θ§≈ΏΧΞ÷Μ”–±≥≤ΩΫαΙΙΕχΟΜ”–ΗΙ≤ΩΫαΙΙΘ§ΕχcactusΘ®catΘ©ΆΜ±δΜα“ΐΤπ≈ΏΧΞΗΙ≤ΩΜ·ΓΘ
Ζ÷ΫΎΜυ“ρΒΡΙΠΡή «Α―≈ΏΧΞΖ÷ΈΣΜ°Ζ÷ΈΣΧεΫΎΘ§Ζ÷ΈΣ»ΐάύΘΚΦδœΕΜυ“ρΘ®gap genesΘ©Θ§≥…Ε‘Μυ“ρΘ®pair-rule genesΘ©ΚΆΧεΫΎΦΪ–‘Μυ“ρΘ®segment polarity geneΘ©ΓΘ’β»ΐΉιΜυ“ρ“≤ «Β»ΦΕΙΊœΒΘ§ΦδœΕΜυ“ρΩΊ÷Τ≥…Ε‘Μυ“ρΘ§≥…Ε‘Μυ“ρΩΊ÷ΤΧεΫΎΦΪ–‘Μυ“ρΓΘ
ΦδœΕΜυ“ρΓΘ»γkrΓΔkniΒ»ΒΡΜυ“ρ≤ζΈοΫΪ≈ΏΖ÷ΈΣœύΒ±”Ύ3ΗωΧεΫΎΒΡ«χ”ρΓΘΦδœΕΜυ“ρ≤ζΈο‘ΎΥϋΟ«ΗςΉ‘ΒΡ±μ¥ο«χΡΎ–Έ≥…≈®Ε»ΧίΕ»Θ§’β–©ΧίΕ»ΧαΙ©ΈΜ÷Ο–≈œΔΗχ≥…Ε‘Μυ“ρΓΘ
≥…Ε‘Μυ“ρΒΡΙΠΡή «Α―ΫΪΦδœΕΜυ“ρΖ÷≥…ΒΡ«χ”ρΫχ“Μ≤ΫΜ°Ζ÷ΈΣΧεΫΎΘ§ «≈ΏΧΞΖ÷ΫΎΒΡ«ΑΉύΓΘ
ΧεΫΎΦΪ–‘Μυ“ρ»γenΓΔwg±Μ≥…Ε‘Μυ“ρΦΛΜνΘ§Ζ÷±π‘ΎΟΩΗωΧεΫΎΒΡ«ΑΓΔΚσ≤ΩœΗΑϊ÷–±μ¥οΘ§“‘–Έ≥…ΚΆΈ§≥÷ΧεΫΎΫαΙΙΓΘ
ΧεΫΎΦΪ–‘Μυ“ρ”÷ΦΛΜνΆ§‘¥“λ–ΈΜυ“ρΘ®homeotic geneΘ§hoxΘ©ΓΘΨωΕ®ΟΩ“ΜΧεΫΎΒΡ–‘÷ ”κ–ΈΧ§ΧΊ’ςΘ§Φ¥―Γ‘ώΧεΫΎœρΡ≥ΗωΖΫœρΖΔ”ΐΓΔΖ÷Μ·Θ§”…”Ύ’βάύΜυ“ρΒΡ≤ζΈοΕΦΚ§”–HDΫαΙΙΘ§Υυ“‘”÷≥ΤΈΣΆ§‘¥“λ–ΈΩρΘ®Κ–Θ©Μυ“ρΘ®homeobox genesΘ§HoxΘ©ΓΘΆ§‘¥“λ–ΈΜυ“ρΒΡΆΜ±δΒΦ÷¬ΖΔ”ΐΒΡ“λ≥ΘΘ§»γ‘Ύ±Ψά¥ΗΟ≥Λ¥ΞΫ«ΒΡΒΊΖΫ≥Λ≥ωΆ»ά¥Θ®ΆΦ14-11Θ©Θ§Εχ‘ΎΗΟ–Έ≥…ΤΫΚβΑτΒΡ≤ΩΈΜ≥Λ≥ωΒΎΕΰΕ‘≥αΘ®ΆΦ14-12Θ©ΓΘ
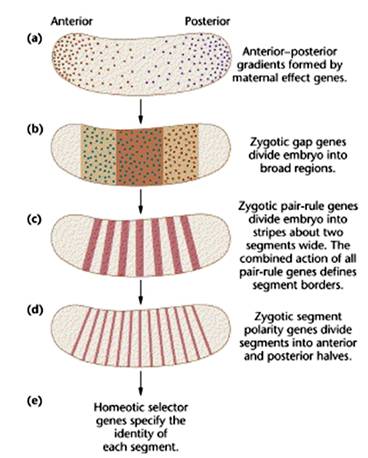
ΆΦ14-10
Ζ÷ΫΎΜυ“ρ

ΆΦ14-11
ΉσΓΔ’ΐ≥ΘΙϊ”§ΒΡ¥ΞΫ«ΈΣΨΏΟΔ¥ΞΫ«Θ§”“ΓΔΆΜ±δΙϊ”§ΒΡ¥ΞΫ«ΖΔ”ΐΈΣΉψ “ΐΉ‘Lehninger Principles of Biochemistry 3rd
ed.

ΆΦ14-12
Ιϊ”§ΒΡΥΪ–ΊΆΜ±δΘ®ΝΫΫΎ÷––ΊΘ§ΟΜ”–Κσ–ΊΘ§“ρΕχΟΜ”–ΤΫΚβΑτΘ©
ΥυΈΫΒΡΉΣ¬Φ“ρΉ”Θ§ «ΡήΙΜ Ε±πΜυ“ρΒΡΤτΕ·Ή”Θ§‘ω«ΩΉ”Θ§≤Δ”κ÷°ΫαΚœΩΊ÷ΤΗΟΜυ“ρΉΣ¬ΦΒΡDNAΫαΚœΒΑΑΉΘ§Ά®ΙΐΕ‘–μΕύΉΣ¬Φ“ρΉ”ΒΡΖ÷ΈωΘ§ΖΔœ÷ΥϋΟ«ΕΦ”–“ΜΗω”κDNAΫαΚœΒΡΧΊ β«χ”ρΘΚ»γΔΌ¬ί–ΐ-ΉΣΫ«-¬ί–ΐΘ®HTHΘ©Θ§ΔΎ–Ω÷ΗΫαΙΙΘ§Δέ¬ί–ΐ-ΜΖ-¬ί–ΐΘ®HLHΘ©Θ§ΔήΆ§‘¥”ρΫαΙΙΘ®DHΘ©÷ς“Σ”…60ΗωΑ±ΜυΥα–ρΝ–ΙΙ≥…Θ§ΗΏΕ»±Θ ΊΘ§ «ΫαΚœDNAΒΡ≤ΩΈΜΘ§”÷≥ΤΈΣΆ§‘¥Κ–Θ®homeoboxΘ©
|