|
在个体发育(ontogeny)中,细胞后代在形态结构和功能上发生差异的过程称为细胞分化(differentiation)。细胞分化不仅发生在胚胎发育中,而是在一生都进行着,以补充衰老和死亡的细胞,如:多能造血干细胞分化为不同血细胞的细胞分化过程。
个体发育是指一个新个体从产生到死亡的全部历程。对两性生殖的生物来说,新个体始于两性配子(精子和卵子)的结合――受精(fertilization)。卵子受精后启动发育程序,形成一个新个体的过程叫做胚胎发育(embryogenesis)。胚胎发育包括受精、卵裂(cleavage)、原肠胚形成(gastrulation)、神经胚形成(neurulation)、器官形成(organogenesis)等几个主要的发育阶段,许多动物还必须经过胚后发育阶段――变态(metamorphosis),才能发育为成体。
第一节
受精与胚胎发育
一、精子和卵的结构
在动物中精母细胞通过减数分裂后形成四个精细胞,精细胞经过精子形成(spermiogenesis)的变态发育过程,排除大部分细胞质,内部发生一系列变化,成为精子。成熟的精子(spermatozoon)形似蝌蚪,分头、尾两部(图14-1)。头内有一个高度浓缩的细胞核,核的前2/3有顶体覆盖(图14-2)。顶体实质上是一个很大的溶酶体,内含多种水解酶,如顶体蛋白酶、透明质酸酶、酸性磷酸酶等。在受精时,精子释放顶体酶消化卵子外面的结构,进入卵内。精于尾部又称鞭毛(flagellum),是精于的运动装置。尾部可分为颈段、中段、主段和末段四部分。颈段很短,其内有两个相互垂直的中心粒。其他三段内的主要结构是由中心粒发出的轴丝,由9+2排列的微管组成。中段短,在轴丝外包有线粒体鞘,为鞭毛的运动提供能量。主段长,没有线粒体鞘,代之以纤维鞘。末段短,仅有轴丝。个别动物如介壳虫的精子没有鞭毛。
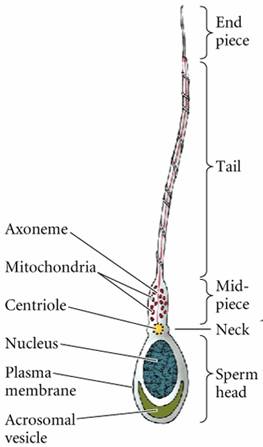
图14-1精子的结构模式图
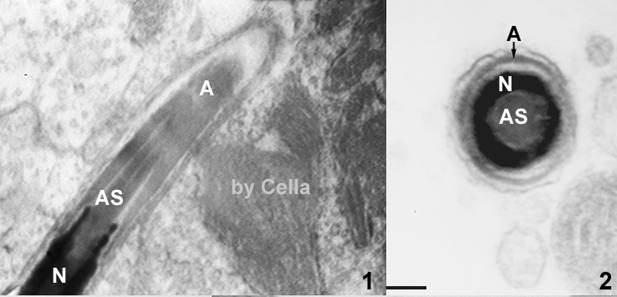
图14-2
绍鸭精子头部的纵切(左)和横切(右)A:顶体;AS:顶体突起;N:细胞核
大多数动物的卵是一个大型的单个细胞,贮存有大量的营养供胚胎发育所用。卵的直径在人和海胆中约为0.1mm,鱼类和两栖类中约为1-2mm,鸟类中可以达到几厘米甚至十几厘米。卵的细胞质中富含蛋白质、脂类和多糖的营养成分称为卵黄(yolk),它们通常存在于卵黄颗粒中,有的种类中卵黄颗粒外还有膜包围。在体外孵化的种类中卵黄的体积可达卵的95%以上。
卵的外面具有外被(coat),其成分主要是糖蛋白,是由卵细胞或其它细胞分泌的。在哺乳动物中这种外被叫做透明带(zona pellucida),其作用是保护卵子,阻止异种精子进入。许多卵的透明带下面(皮质部,cortex)还有一层分泌性的囊泡,称为皮层颗粒(cortical granules),受精时以外排的方式释放皮层颗粒能引起透明带结构变化,形成受精膜,阻止其它精子进入。
卵细胞实际上是停留在第一次或第二次减数分裂阶段的卵母细胞,直到受精以后,才完成减数分裂,并释放极体。人类从卵巢排出的卵子处于第二次成熟分裂的中期,在受精时才完成第二次分裂。若未受精,则于排卵后12-24小时退化。
二、受精
受精包括精子的获能(capacitation)、顶体反应(acrosome
reaction)、皮层反应(cortical reaction)、原核形成和融合等过程(图14-3)。
刚排出的精子虽有运动能力,但不能穿过卵子的放射冠和透明带,只有接受雌性生殖道的分泌物(获能因子),才具有受精能力,这种作用称为精子获能。在体外条件下,血清白蛋白(serum albumin)、高密脂蛋白(high-density lipoproteins)、氨基聚糖(glycosaminoglycans)、孕酮、钙离子载体(A23187)、ZP3、CO2等均可促进精子获能。获能后精子表面阻止受精的附睾蛋白和精浆蛋白被除去或改变,质膜胆固醇流失,与卵子结合的受体暴露。获能期间,钙通道被激活、耗氧量和糖酵解明显增加,pH升高,腺苷酸环化酶激活,导致胞内cAMP含量升高,蛋白激酶A活化,精子活力增加,顶体酶原转化为有活性的顶体酶。
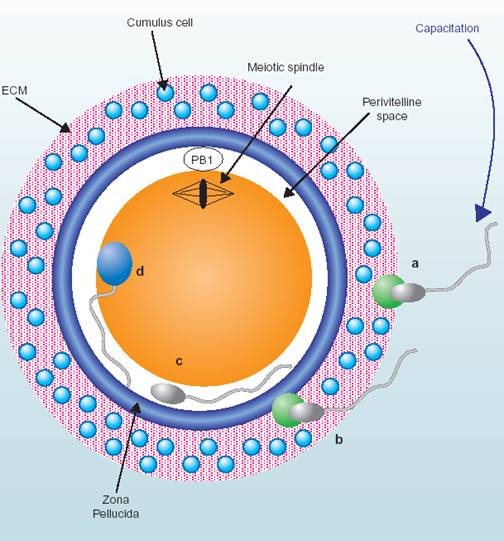
图14-3
受精的早期事件 引自Janice P. Evans 2002
获能的精子首先借助头后部(顶体后区)质膜PH-20的透明质酸酶活性穿过卵母细胞外卵丘细胞层,到达透明带,然后精子通过头前部(顶体区)质膜表面的ZP结合蛋白与卵母细胞透明带发生识别,完成结合过程,诱发顶体反应,即:顶体以外排的方式释放出水解酶,将卵子的透明带和卵黄膜溶解,形成精子穿过的通道。在海胆中此时精子的前端在微丝的作用下还会形成顶体突起(哺乳动物不具有)。
精子进入卵子后,导致卵膜去极化,动作电位由受精点传至膜表面,使精子受体失活。在精子接触点上,磷脂酰肌醇信号途径启动,产生DAG和IP3。DAG激活蛋白激酶C,启动DNA的复制。IP3导致内质网中的钙离子释放,钙浓度呈有规律的跃迁,称为钙波。钙信号进而引起皮层反应,即:卵细胞膜下的皮层颗粒(内含酶类的囊泡),以外排的方式,进入卵黄膜和卵膜之间的腔隙,酶类引起透明带“硬化”,形成受精膜。
精子的刺激使处于休眠状态的卵子被激活,重新回到减数分裂阶段,迅速完成第二次分裂,释放极体。此时精子和卵子的细胞核分别称为雄原核(male pronucleus)和雌原核(female pronucleus)。两个原核逐渐在细胞中部靠拢,核膜随即消失,染色体混合,形成二倍体的受精卵(fertilized ovum),又称合子(zygote)。
通常一个卵子和一个精子结合,多精进入会形成多余的分裂极和纺锤体,导致细胞异常分裂而使胚胎发育终止。受精后通过两种机制阻止多精进入,一是膜瞬间去极化;二是通过皮层反应,破坏精子受体和形成受精膜。受精后精子的核、中心粒(鞭毛基粒)和线粒体均注入卵中。线粒体之所以表现为母系遗传,是因为受精卵中只有仅母本线粒体可以存活。
许多昆虫、软体动物、软骨鱼、有尾两栖类、爬行类和鸟类在受精时有多于一个精子进入卵内,但只有一个雄原核与雌原核发生融合,参与发育,其余的精核逐渐退化消失。有时多余的精于也发生一些畸形的分裂,但不妨碍正常卵裂和发育。这种多精受精称为生理性多精受精。
三、精卵识别
自然环境中,几种海胆的精子可以接触一个卵子,产生顶体反应,但异种精子不能与卵子融合,这是因为精子表面的结合素能与卵细胞膜上特异的受体结合,而达到同种识别的目的。
在哺乳动物小鼠中,精卵的识别同样涉及表面糖蛋白之间的相互作用,通常把卵子透明带中与精子结合的蛋白(ZP)叫做精子受体,而把精子表面与ZP结合的蛋白叫做卵结合蛋白。
小鼠透明带中有三种糖蛋白,命名为ZP1、ZP2、ZP3,分子量分别为200、120和83KD。ZP2和ZP3形成异二聚体,ZP1与二聚体交联,形成三维网状结构。ZP3是第一精子受体,能诱发顶体反应。用ZP3蛋白处理精子,则失去受精能力。
β1,4-半乳糖转移酶-I(β1,4-galactosyltransferase,GalTase)是精子表面负责与ZP3结合的蛋白(图14-4)。GalTase-I能结合ZP3糖链末端的N-乙酰葡糖胺(N-acetylglucosamine)残基,实验证明纯化的GalTase-I及其抗体均抑制精卵结合。GalTase-I能激活G蛋白,引起精子顶体反应,GalTase-I基因敲除的小鼠其精子缺乏顶体反应和穿越透明带的能力。
精子穿过透明带,到达卵周隙后位于精子头后部质膜的受精素(fertilin,也叫ADAM)与卵母细胞质膜上的整合素发生识别和结合(图14-5),并启动精子与卵母细胞的质膜融合系统,最终实现受精过程。
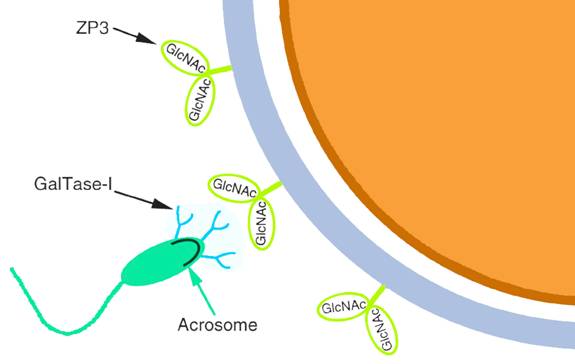
图14-4
ZP3与GalTase 引自D. J. Miller 2000
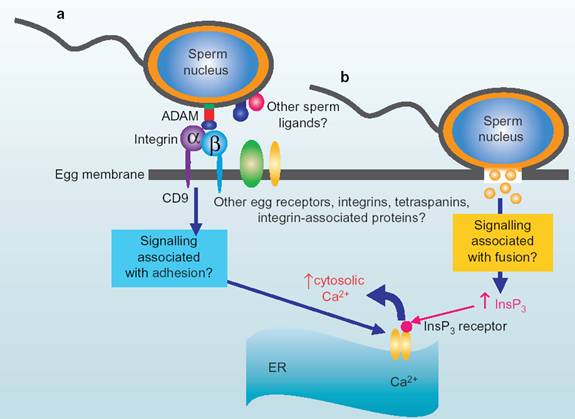
图14-5精子和卵子细胞膜的相互作用,引自Janice P. Evans 2002
四、卵裂与胚胎发育
卵裂(cleavage)是指受精卵经过分裂,将卵质分配到子细胞的过程,分裂产生的细胞叫做分裂球(blastomere)。卵裂和一般有丝分裂相似,但不经过间期,所以卵裂期间仅仅是细胞数目的增加,不伴随着细胞生长。随着细胞数量增加,子细胞的核质比逐渐增大,直到接近正常核质比时,分裂球才开始生长,进入到一般的有丝分裂过程。
卵裂的方式与卵黄含量和分布有关,较大、卵黄较多的卵是以偏裂的方式进行的(如昆虫的表面卵裂),反之多以均裂的方式进行(如哺乳动物的卵)。分裂面与卵的极轴一致的卵裂称为经裂(meridional cleavage),与极轴垂直的称为纬裂(latitudinal cleavage)。卵裂方向和纺锤体的方向有关,细胞质成分和环境信号均可以影响纺锤体的方向。
两栖类的卵受精后两小时就开始卵裂,第1、2次为经裂,第3次为纬裂,不对称,在动物极形成4个较小的细胞,在植物极形成4个较大的细胞。
哺乳动物的卵裂较慢,受精1天后才开始卵裂,8细胞之前,分裂球之间结合比较松散,8细胞之后突然紧密化(compaction),即:通过细胞连接形成致密的球体。16细胞期,内部1-2个细胞属于内细胞团,将来发育为胚胎,而其外周细胞变为滋养细胞,不参与组成胚胎结构,而是参与形成绒毛膜。
通常动物的胚胎在64细胞以前为实心体,称为桑椹胚,在128细胞阶段,细胞团内部空隙扩大,成为充满液体的囊胚腔(blastocoel),此时的胚胎称为囊胚。
囊胚继续发育,部分位于外表面的细胞通过各种细胞运动方式(如:移入、内卷、内陷)进入内部从而形成一个二层或三层的原肠胚。这种细胞迁移运动过程称为原肠形成(gastrulation)。留在外面的称为外胚层(ectoderm),迁移到里面的称为内胚层(endoderm)或中、内胚层(meso-endoderm)。原来的囊胚腔亦随原肠腔的形成而逐渐消失。
三胚层形成的组织和器官有:外胚层形成神经系统、表皮、皮肤腺、毛发、指甲、爪和牙齿等;中胚层形成骨骼、肌肉、泌尿生殖系统、淋巴组织、结缔组织、血液;内胚层形成呼吸系统、消化道、肝、胰。
|