|
第三节 线粒体与叶绿体的蛋白质定向转运
线粒体的蛋白合成能力有限,大量线粒体蛋白在细胞质中合成,定向转运到线粒体。这些蛋白质在在运输以前,以未折叠的前体形式存在,与之结合的分子伴娘(属hsp70家族)保持前体蛋白质处于非折叠状态。通常前体蛋白N端有一段信号序列称为导肽、前导肽或转运肽(leader sequence、presequence或transit-peptide),完成转运后被信号肽酶(signal peptidase)切除,就成为成熟蛋白,这种现象就叫做后转译(posttranslation,图7-27)。
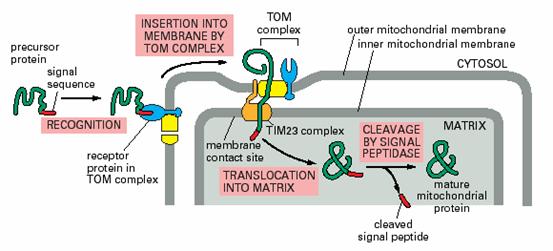
图7-27 线粒体蛋白质的定向转运 引自Molecular
Biology of the Cell. 4th ed. 2002
线粒体前体蛋白信号序列的特点是:①多位于肽链的N端,由大约20个氨基酸构成;②没有带负电荷的氨基酸,形成一个两性α螺旋,带正电荷的氨基酸残基和不带电荷的疏水氨基酸残基分别位于螺旋的两侧,现在认为这个螺旋与转位因子的识别有关;③对所牵引的蛋白质没有特异性要求,非线粒体蛋白连接上此类信号序列,也会被转运到线粒体。此外有些信号序列位于蛋白质内部,完成转运后不被切除,还有些信号序列位于前体蛋白C端,如线粒体的DNA解旋酶 Hmil。
表7-1 线粒体蛋白分选信号*
|
信号序列
|
定位
|
转运装置
|
信号序列位置
|
|
位于N端,富含带正电荷的和疏水的氨基酸,形成两性α螺旋,完成转运后被切除。
|
基质
|
TOM
TIM23
|

|
|
不被切除,含疏水性的停止转移序列,被安插到外膜。
|
外膜
|
TOM
|

|
|
被切除,含疏水性的停止转移序列,被安插到内膜。
|
内膜
|
TOM
TIM23
|

|
|
含两个信号序列,首先转运到基质,第一个信号序列被切除,第二个信号序列引导蛋白进入内膜或膜间隙。
|
内膜
膜间隙
|
TOM
TIM23
|
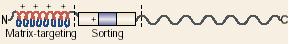
|
|
结构类似于N端信号序列,但位于蛋白质内部。
|
内膜
|
TOM
TIM23
|

|
|
为线粒体代谢物的转运蛋白,如腺苷转位酶,具有多个内部信号序列和停止转移序列,形成多次跨膜蛋白。
|
内膜
|
TOM
TIM22
|

|
*根据N. Pfanner and A. Geissler 2001改编
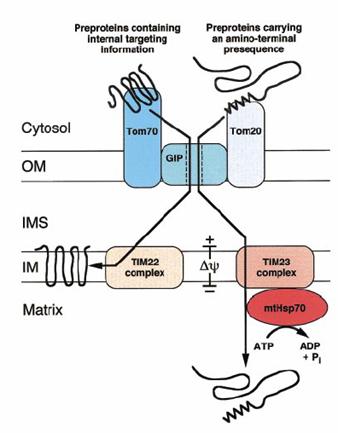
图7-28 线粒体的蛋白转运装置——TOM和TIM复合体 引自Joachim Rassow and Nikolaus Pfanner 2000
蛋白质的转运涉及多种蛋白复合体,即转位因子(translocator,图7-28),由两部分构成的:受体和蛋白质通过的孔道。主要包括:①TOM复合体,负责通过外膜,进入膜间隙,在酵母中TOM70负责转运内部具有信号序列的蛋白,TOM20负责转运N端具有信号序列的蛋白,这两种蛋白的功能都相当于内质网上的SPR受体,在人类线粒体中hTom34的功能与TOM70相当。TOM复合体的通道被称为GIP(general import pore),就相当于内质网上的SEC61复合体,主要由Tom40构成, 还包括Tom22, Tom7, Tom6和Tom5;②TIM复合体,其中TIM23负责将蛋白质转运到基质,也可将某些蛋白质安插在内膜;TIM22负责将线粒体的代谢物运输蛋白,如ADP/ATP和磷酸的转运蛋白插入内膜;③OXA复合体:负责将线粒体自身合成的蛋白质插到内膜上,同样也可使经由TOM/TIM复合体进入基质的蛋白质插入内膜。
线粒体具有四个功能区隔,即外膜、内膜、膜间隙、基质。进入不同部位的蛋白具有不同的转运途径。
进入外膜的蛋白具有不被切除的N端信号序列,其后还有疏水性序列作为停止转移序列,然后蛋白质被TOM复合体安装到外膜上,如线粒体的各类孔蛋白。
进入基质蛋白质可以先通过TOM复合体进入膜间隙,然后通过TIM复合体进入基质。也可以通过线粒体内、外膜间的接触点(鼠肝直径1um线粒体上约115个接触点),一步进入基质,在接触点上TOM与TIM协同作用完成蛋白质向基质的输入(图7-29)。
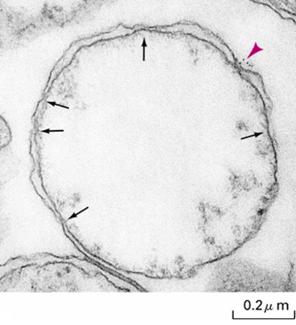
图7-29 线粒体内外膜的接触点
进入线粒体内膜和膜间隙的蛋白具有以下几种情况(图7-30):①蛋白N端具有两个信序列,首先被运送到基质,然后N端信号肽被切除,暴露出导向内膜的信号序列,在OXA的帮助下插入内膜。如果第二段信号序列被内膜外表面的异二聚体内膜蛋白酶(heterodimeric inner
membrane peptidase,Imp1/imp2)切除,则成为膜间隙蛋白。②N端信号序列的后面有一段疏水序列,扮演停止转移序列的角色,能与TIM23复合体结合,当进入基质的信号序列被切除后,脱离转位因子复合体而进入内膜,如果插入膜中的部分又被酶切除,侧成为定位于膜间隙的蛋白。③线粒体内膜上负责代谢底物/产物转运的蛋白,如腺苷转位酶是多次跨膜蛋白,其N端没有可被切除的信号序列,但包含3-6个内部信号序列,可被TIM22复合体插到内膜上。
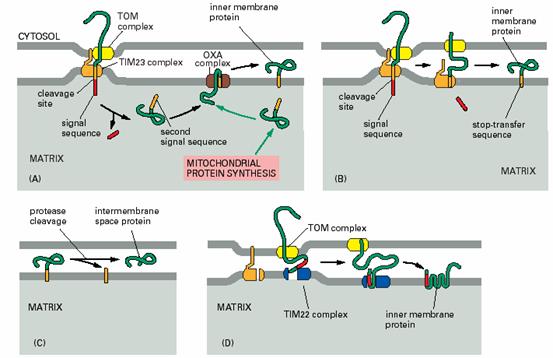
图7-30 蛋白质进入内膜和膜间隙 引自Molecular
Biology of the Cell. 4th ed. 2002,(A. 进入内膜和膜间隙的前体蛋白具有两个信号序列,经TOM/TIM23进入基质后,第二个信号序列使蛋白通过OXA复合体被安插到内膜上;B. 进入内膜和膜间隙的前体蛋白信号序列后具有停止转移序列,被TIM23安插在膜上,C. 通过途径A、B插入内膜的蛋白,被位于内膜的蛋白酶加工,成为膜间隙的可溶性蛋白,D. 线粒体代谢物的转运器为多次跨膜蛋白,被TIM22安插到内膜中)
蛋白质的输入是一个耗能的过程,能量的来源为水解ATP和利用质子动力势.能量消耗在线粒体外和进入线粒体基质两步上,在线粒体外解除与前体蛋白质结合的分子伴娘,需要通过水解ATP获得能量;在通过TIM复合体进入基质时利用质子动力势作为动力。虽然目前含不清楚质子动力势是如何被利用的,但解偶连接如DNP能抑制蛋白质的转运。前体蛋白进入线粒体基质后,线粒体hsp70一个接一个的结合在蛋白质线性分子上,像齿轮一样将蛋白质“铰进(hand over hand)”基质,这一过程也需要消耗ATP。然后线粒体hsp70将蛋白质交给hsp60,完成折叠。
叶绿体大约含2000-2500 种蛋白,叶绿体基因组编码的不足100 种,因此大量的蛋白质也是由核基因编码,在细胞质中合成,然后定向转运到叶绿体。
叶绿体蛋白的转运机理与线粒体的相似。如:通常前体蛋白N端具有信号序列,完成转运后被信号肽酶切除,和线粒体一样发生后转译;每一种膜上有特定的转位因子;内外膜具有接触点(contact site);需要能量,同样利用ATP和质子动力势。但是两者的蛋白质转运体系中除了某些hsp分子相同外,转位因子复合体是不同的。叶绿体外膜的转位因子被称为TOC复合体,内膜的转位因子被称为TIC复合体。
叶绿体有6个区隔:①外膜、②膜间隙、③内膜、④基质、⑤类囊体膜、⑥类囊体腔。因此叶绿体的蛋白转运途径更复杂一些。
叶绿体前体蛋白的N端信号序列长度为20-150 个氨基酸残基,同一植物不同前体蛋白的N端序列不具有同源性,分为3 个部分:N 端缺乏带正电荷的氨基酸,以及甘氨酸和脯氨酸;C 端形成两性β折叠;中间富含羟基化的氨基酸,如丝氨酸和苏氨酸。
转运到叶基质的前体蛋白具有典型的N端序列。转运到叶绿体内膜和类囊体的前体蛋白含有两个N端信号序列,第一个被切除后,暴露出第二个信号序列,将蛋白导向内膜或类囊体膜(图7-30)。转运到叶绿体外膜上的蛋白分两类:一类含内部信号序列;另一类蛋白的N 端序列被切除后,其后的停止转移序列使蛋白质定位在外膜。
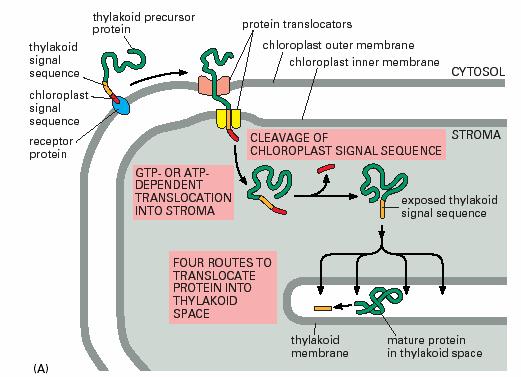
图7-31 叶绿体的蛋白质定向转运 引自Molecular
Biology of the Cell. 4th ed. 2002
|