|
第四节 细胞周期调控
一、研究背景
Rao和Johnson(1970、1972、1974)将Hela细胞同步于不同阶段,然后与M期细胞混合,在灭活仙台病毒介导下,诱导细胞融合,发现与M期细胞融合的间期细胞产生了形态各异的早熟凝集染色体(prematurely condensed chromosome,PCC),这种现象叫做早熟染色体凝集(premature chromosome condensation)。
G1期PCC为单线状,因DNA未复制。
S期PCC为粉末状,因DNA由多个部位开始复制。
G2期PCC为双线染色体,说明DNA复制已完成。
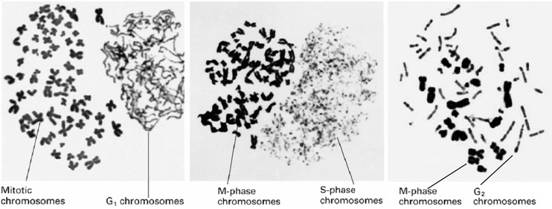
图13-15 不同形态的PCC
不仅同类M期细胞可以诱导PCC,不同类的M期细胞也可以诱导PCC产生,如人和蟾蜍的细胞融合时同样有这种效果,这就意味着M期细胞具有某种促进间期细胞进行分裂的因子,即成熟促进因子(maturation promoting factor,MPF)。
早在1960s,Yoshio Masui发现成熟蛙卵的提取物能促进未成熟卵的胚胞破裂(Germinal Vesicle Breakdown,GVBD),后来Sunkara将不同时期Hela细胞的提取液注射到蛙卵母细胞中,发现G1和S期的抽取物不能诱导GVBD,而G2和M期的则具有促进胚胞破裂的功能,它将这种诱导物质称为有丝分裂因子(MF)。后来在CHO细胞,酵母和粘菌中也提取出相同性质的MF。这类物质被统称为MPF。
1960s Leland Hartwell以芽殖酵母(图13-16)为实验材料,利用阻断在不同细胞周期阶段的温度敏感突变株(在适宜的温度下和野生型一样),分离出了几十个与细胞分裂有关的基因(cell division cycle gene,CDC)。如芽殖酵母的cdc28基因,在G2/M转换点发挥重要的功能。Hartwell还通过研究酵母菌细胞对放射线的感受性,提出了checkpoint(细胞周期检验点)的概念,意指当DNA受到损伤时,细胞周期会停下来。
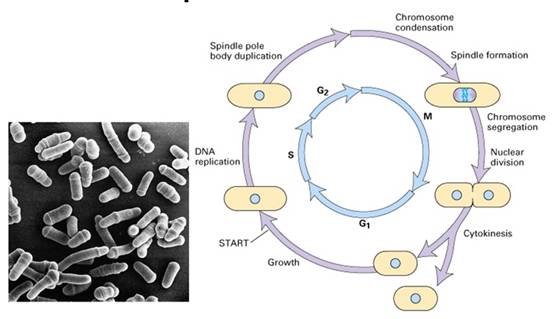
图13-16 裂殖酵母细胞周期
1970s Paul Nurse等人以裂殖酵母(图13-17)为实验材料,同样发现了许多细胞周期调控基因,如:裂殖酵母cdc2、cdc25的突变型和在限制的温度下无法分裂;wee1突变型则提早分裂,而cdc25和wee1都发生突变的个体却会正常地分裂(图13-18)。进一步的研究发现cdc2和cdc28都编码一个34KD的蛋白激酶,促进细胞周期的进行。而weel和cdc25分别表现为抑制和促进CDC2的活性。这也解释了为何cdc25和wee1双重突变的个体可以恢复野生型的表型。
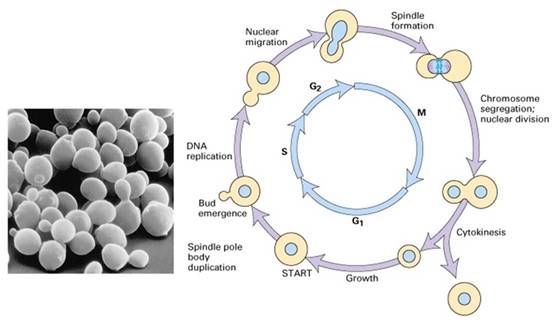
图13-17 芽殖酵母细胞周期
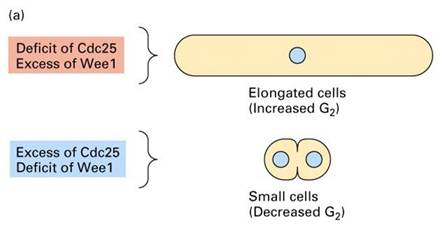
图13-18
Cdc25表达不足,细胞长得过长而不分裂;Wee1表达不足,细胞很小就开始分裂了
1983年Timothy Hunt首次发现海胆卵受精后,在其卵裂过程中两种蛋白质的含量随细胞周期剧烈振荡,在每一轮间期开始合成,G2/M时达到高峰,M结束后突然消失,下轮间期又重新合成,故命名为周期蛋白(cyclin)。后来在青蛙、爪蟾、海胆、果蝇和酵母中均发现类似的情况,各类动物来源的细胞周期蛋白mRNA均能诱导蛙卵的成熟。用海洋无脊椎动物和两栖类的卵为实验材料进行这类实验,好处在于卵的量比较大,而且在胚胎发育的早期,细胞分裂是同步化的。
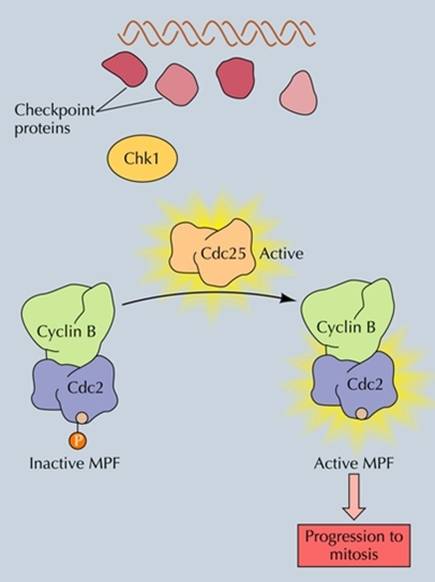
图13-19 MPF=CDC2+Cyclin B
1988年M. J. Lohka 纯化了爪蟾的MPF,经鉴定由32KD和45KD两种蛋白组成,二者结合可使多种蛋白质磷酸化(图13-19)。后来Paul Nurse(1990)进一步的实验证明P32实际上是CDC2的同源物,而P45是cyclinB的同源物,从而将细胞周期三个领域的研究联系在一起。2001年10月8日美国人Leland Hartwell、英国人Paul Nurse、Timothy Hunt因对细胞周期调控机理的研究而荣获诺贝尔生理医学奖(图13-20)。

图13-20 2001年诺贝尔生理医学奖获得者(图片来自http://www.nobel.se/)
二、CDK
CDC2与细胞周期蛋白结合才具有激酶的活性,称为细胞周期蛋白依赖性激酶(cyclin-dependent kinase,CDK),因此CDC2又被称为CDK1,激活的CDK1可将靶蛋白磷酸化而产生相应的生理效应,如将核纤层蛋白磷酸化导致核纤层解体、核膜消失,将H1磷酸化导致染色体的凝缩等等。这些效应的最终结果是细胞周期的不断运行。因此,CDK激酶和其调节因子又被称作细胞周期引擎。
目前发现的CDK在动物中有7种。各种CDK分子均含有一段相似的激酶结构域,这一区域有一段保守序列,即PSTAIRE,与周期蛋白的结合有关。
三、CKI
细胞中还具有细胞周期蛋白依赖性激酶抑制因子(CDK inhibitor,CKI)对细胞周期起负调控作用,目前发现的CKI分为两大家族:
①Ink4(Inhibitor of cdk 4),如P16ink4a、P15ink4b、P18ink4c、P19ink4d,特异性抑制cdk4・cyclin D1、cdk6・cyclin D1复合物。
②Kip(Kinase inhibition protein):包括P21cip1 (cyclin
inhibition protein 1)、P27kip1(kinase
inhibition protein 1)、P57kip2等,能抑制大多数CDK的激酶活性,P21cip1还能与DNA聚合酶δ的辅助因子PCNA(proliferating cell nuclear antigen)结合,直接抑制DNA的合成(图13-21)。
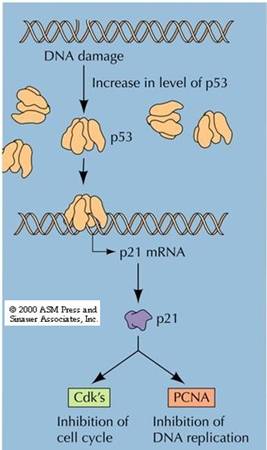
图13-21 P21cip1抑制CDK和PCNA
四、Cyclin
周期蛋白不仅仅起激活CDK的作用,还决定了CDK何时、何处、将何种底物磷酸化,从而推动细胞周期的前进。目前从芽殖酵母、裂殖酵母和各类动物中分离出的周期蛋白有30余种,在脊椎动物中为A1-2、B1-3 、C、 D1-3、E1-2、F、G、H等。分为G1型、G1/S型S型和M型4类(见表13-1)。各类周期蛋白均含有一段约100个氨基酸的保守序列,称为周期蛋白框,介导周期蛋白与CDK结合。
表13-1不同类型的周期蛋白
|
激酶复合体 |
脊椎动物 |
芽殖酵母 |
|
Cyclin |
CDK |
Cyclin |
CDK |
|
G1-CDK |
Cyclin D* |
CDK4 、6 |
Cln 3 |
CDK1(CDC28) |
|
G1/S-CDK |
Cyclin E |
CDK2 |
Cln 1、2 |
CDK1(CDC28) |
|
S-CDK |
Cyclin A |
CDK2 |
Clb 5、6 |
CDK1(CDC28) |
|
M-CDK |
Cyclin B |
CDK1(CDC2) |
Clb 1-4 |
CDK1(CDC28) |
*包括D1-3,各亚型cyclin
D,在不同细胞中的表达量不同,但具有相同的功效
细胞在生长因子的刺激下,G1期cyclin D表达,并与CDK4、CDK6结合,使下游的蛋白质如Rb磷酸化,磷酸化的Rb释放出转录因子E2F,促进许多基因的转录,如编码cyclinE、A和CDK1的基因。
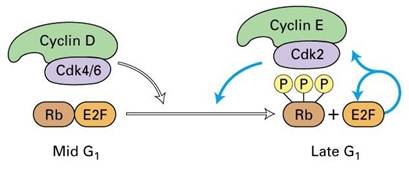
图13-22 Cyclin D与CDK结合使Rb释放结合的转录因子E2F
在G1-S期, cyclinE与CDK2结合,促进细胞通过G1/S限制点而进入S期。向细胞内注射CyclinE的抗体能使细胞停滞于G1期,说明细胞进入S期需要CyclinE的参与。同样将CyclinA的抗体注射到细胞内,发现能抑制细胞的DNA合成,推测CyclinA是DNA复制所必需的。
在G2-M期,cyclinA、cyclinB与CDK1结合,CDK1使底物蛋白磷酸化、如将组蛋白H1磷酸化导致染色体凝缩,核纤层蛋白磷酸化使核膜解体等下游细胞周期事件。
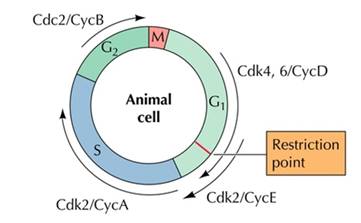
图13-23 Cyclin的周期性变化
在中期当MPF活性达到最高时,通过一种未知的途径,激活后期促进因子APC,将泛素连接在cyclinB上,导致cyclinB被蛋白酶体(proteasome)降解,完成一个细胞周期(图13-24)。
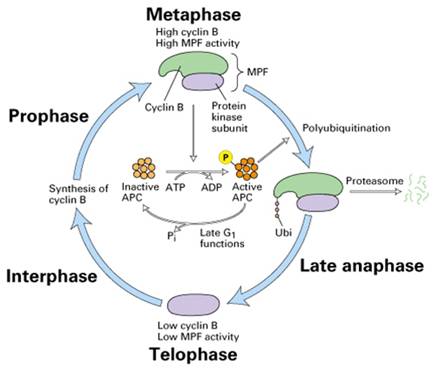
图13-24 Cyclin B的降解途径
分裂期周期蛋白N端有一段序列与其降解有关,称降解盒(destruction box,图13-25)。当MPF活性达到最高时,通过泛素连接酶催化泛素与cyclin结合,cyclin随之被26S蛋白酶体水解。G1周期蛋白也通过类似的途径降解,但其N端没有降解盒,C端有一段PEST序列与其降解有关。
泛素由76个氨基酸组成,高度保守,普遍存在于真核细胞,故名泛素。共价结合泛素的蛋白质能被蛋白酶体识别和降解,这是细胞内短寿命蛋白和一些异常蛋白降解的普遍途径,泛素相当于蛋白质被摧毁的标签。26S蛋白酶体是一个大型的蛋白酶,可将泛素化的蛋白质分解成短肽。
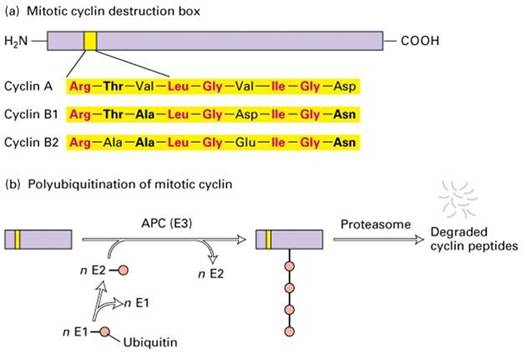
图13-25 细胞周期蛋白的降解盒与降解途径
在蛋白质的泛素化过程中(图13-25),E1(ubiquitin-activating enzyme,泛素激活酶)水解ATP获取能量,通过其活性位置的半胱氨酸残基与泛素的羧基末端形成高能硫酯键而激活泛素,然后E1将泛素交给E2(ubiquitin-conjugating enzyme,泛素结合酶),最后在E3(ubiquitin-ligase,泛素连接酶)的作用下将泛素转移到靶蛋白上。参与细胞周期调控的泛素连接酶至少有两类,其中SCF(skp1-cullin-F-box protein,三个蛋白构成的复合体)负责将泛素连接到G1/S期周期蛋白和某些CKI上,APC(anaphase promoting complex)负责将泛素连接到M期周期蛋白上。
五、DNA复制当且仅当一次
DNA的复制是由起始复制点(origins of replication)开始的,起始复制点也就是上一章提到的自主复制序列,散布在染色体上。在整过细胞周期中,起始复制点上结合有起始识别复合体(Origin recognition complex,
ORC),其作用就象一个停泊点,供其它调节因子停靠。
CDC6是其中的一个调节因子,在G1期CDC6含量瞬间提高,CDC6结合在ORC上,在ATP供能下,促进6个亚单位构成的MCM复合体和其他一些蛋白结合到ORC上,形成前复制复合体(pre-replicative complex,pre-RC),MCM实际上就是DNA解旋酶(helicase)。
S-CDK触发pre-RC的启动,同时阻止了DNA再次进行复制,因为S-CDK将CDC6磷酸化,使其脱离ORC,磷酸化的CDC6随后被SCF参与的泛素化途径降解;S-CDK还可以将某些MCM磷酸化,使其被输出细胞核。其它一些CDK也参与阻止pre-RC的再次形成,从而保证了DNA的复制当且仅当一次(图13-26)。
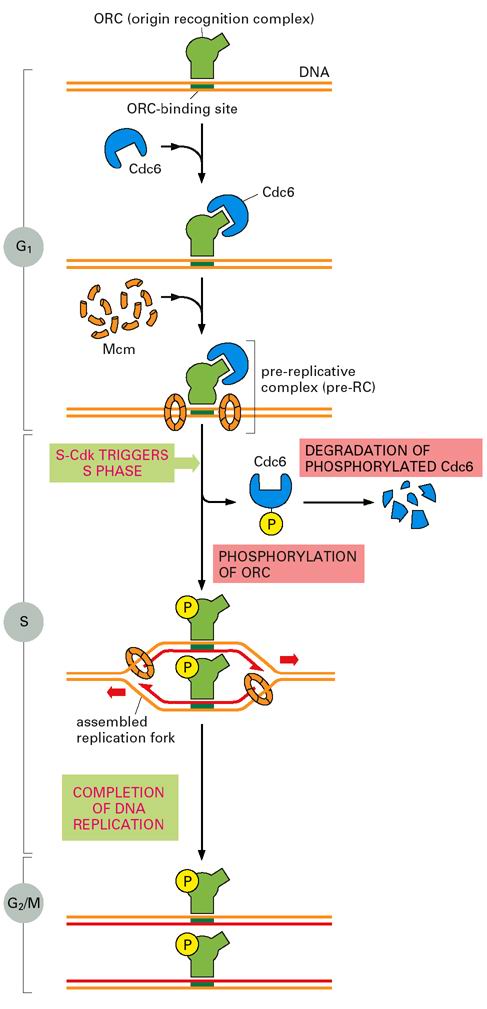
图13-26
每个细胞周期启动一次DNA复制(图片来自Molecular Biology of the Cell 4th
ed.)
六、M期CDK的激活
M期CDK的激活起始于分裂期cyclin的积累,在胚胎细胞周期中cyclin一直在合成,其浓度决定于降解的速度;但在大多数细胞的有丝分裂周期中,cyclin的积累是因为在G2-M期M-cyclin基因转录的增强。
随着M-cyclin的积累,结合周期蛋白的M-CDK(CDK1)增加,但是没有活性,这是因为Wee1激酶将CDK1的Thr14和Tyr15磷酸化的缘故,这种机制保证了CDK-cyclin能够不断积累,然后在需要的时候突然释放。
在M期,一方面Wee1的活性下降,另一方面CDC25使CDK去磷酸化,去除了CDK活化的障碍。CDC25可被两种激酶激活,一是polo激酶,另一个是M-CDK本身。激活的M-CDK还可以抑制它的抑制因子Wee1的活性,形成一个反馈环。因此不难想象只要有少量的CDK被CDC25或polo激活,立即就会有大量的CDK被活化。
CDK的激活还需要Thr161的磷酸化,它是在CDK激酶(CDK activating kinase CAK)的作用下完成的(图13-27)。
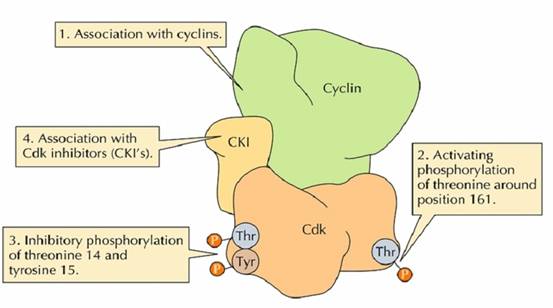
图13-27 CDK1的激活需要Thr14和Tyr15去磷酸化和Tyr161的磷酸化
七、细胞周期检验点
细胞要分裂,必须正确复制DNA和达到一定的体积,在获得足够物质支持分裂以前,细胞不可能进行分裂。细胞周期的运行,是在一系列称为检验点(check point)的严格检控下进行的,当DNA发生损伤,复制不完全或纺锤体形成不正常,周期将被阻断。
细胞周期检验点由感受异常事件的感受器、信号传导通路和效应器构成,主要检验点包括(图13-28):
G1/S检验点:在酵母中称start点,在哺乳动物中称R点(restriction point),控制细胞由静止状态的G1进入DNA合成期,相关的事件包括:DNA是否损伤?细胞外环境是否适宜?细胞体积是否足够大?
S期检验点:DNA复制是否完成?
G2/M检验点:是决定细胞一分为二的控制点,相关的事件包括:DNA是否损伤?细胞体积是否足够大?
中-后期检验点(纺锤体组装检验点):任何一个着丝点没有正确连接到纺锤体上,都会抑制APC的活性,引起细胞周期中断。
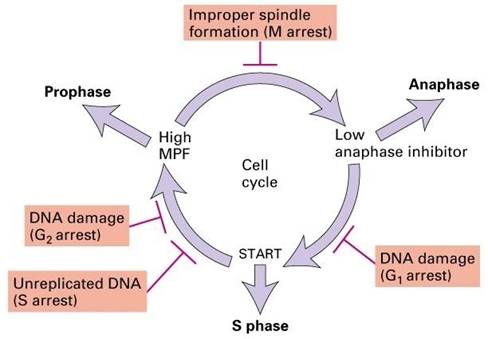
图13-28 四个主要的检验点
ATM(ataxia telangiectasia-mutated gene)是与DNA损伤检验有关的一个重要基因。最早发现于毛细血管扩张性共济失调症患者,人类中大约有1%的人是ATM缺失的杂合子,表现出对电离辐射敏感和易患癌症。正常细胞经放射处理后,DNA损伤会激活修复机制,如DNA不能修复则诱导细胞凋亡,总之不会形成变异的细胞。
ATM编码一个蛋白激酶,结合在损伤的DNA上,能将某些蛋白磷酸化,中断细胞周期。其信号通路有两条。
一条是激活Chk1(checkpoint kinase),Chk1引起CDC25的Ser216磷酸化,通过抑制CDC25的活性,抑制M-CDK的活性,使细胞周期中断。
另一条是激活Chk2,使P53被磷酸化而激活,然后P53作为转录因子,导致P21的表达,P21抑制G1-S期CDK的活性,从而使细胞周期阻断。
八、生长因子对细胞增殖的影响
单细胞生物的增值取决于营养是否足够,多细胞生物细胞的增值取决于机体是否需要。这种需要是通过细胞通信来实现的。
生长因子是一大类与细胞增殖有关的信号物质,目前发现的生长因子多达几十种,多数有促进细胞增殖的功能,故又称有丝分裂原(mitogen),如表皮生长因子(EGF)、神经生长因子(NGF),少数具有抑制作用如抑素(chalone),肿瘤坏死因子(TNF),个别如转化生长因子β(TGF-β)具有双重调节作用,能促进一类细胞的增值,而抑制另一类细胞。
生长因子不由特定腺体产生,主要通过旁分泌作用于邻近细胞。各种生长因子分子量大小不同,如:肝细胞生长因子(HGF)由674个氨基酸组成,分子量达80KD,内皮素仅由21个氨基酸组成。大多数生长因子仅由一条肽链组成,如EGF、TGF-α、FGF,而PDGF、NGF、TGF-β,肝细胞生长因子HGF由两条肽组成。
生长因子的信号通路主要有:ras途径,cAMP途径和磷脂酰肌醇途径。如通过ras途径,激活MAPK,MAPK进入细胞核内,促进细胞增殖相关基因的表达。如通过一种未知的途径激活c-myc,myc作为转录因子促进cyclin D、SCF、E2F等G1-S有关的许多基因表达,细胞进入G1期(图13-29)。
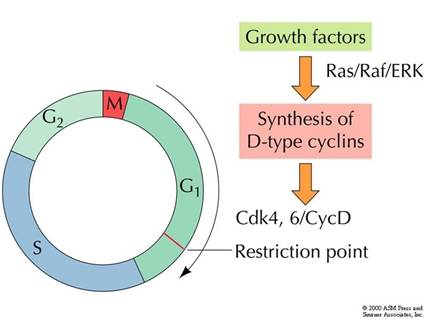
图13-29
生长因子的作用机理
图13-30 细胞周期和其他的一些细胞生理过程就像多米诺骨牌
|