第三节 核仁
核仁(nucleolus)见于间期的细胞核内,呈圆球形,一般1-2个,也有多达3-5个的。核仁的位置不固定,或位于核中央,或靠近内核膜,核仁的数量和大小因细胞种类和功能而异。一般蛋白质合成旺盛和分裂增殖较快的细胞有较大和数目较多的核仁,反之核仁很小或缺如。核仁在分裂前期消失,分裂末期又重新出现。核仁的主要功能是转录rRNA和组装核糖体单位。
一、
核仁形态
核仁与其他的细胞器不同,周围没有界膜包围,在电子显微镜下可辨认出有3个特征性的区域(图12-28):①纤维中心(fibrillar centers,FC):是被致密纤维包围的一个或几个低电子密度的圆形结构,主要成分为RNA聚合酶和rDNA,这些rDNA是裸露的分子。②致密纤维组分(dense fibrillar component,DFC):呈环形或半月形包围FC,由致密的纤维构成,是新合成的RNP(指结合蛋白质的rRNA),转录主要发生在FC与DFC的交界处。③颗粒组分(granular component,GC):由直径15-20nm的颗粒构成,是不同加工阶段的RNP。
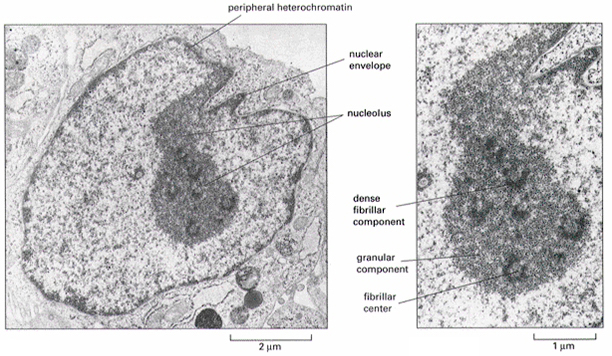
图12-28
核仁结构
核仁相随染色质分为两部分,一部分位于核仁周围,称为核仁周染色质,属异染色质,一部分位于核仁内,为常染色质,即核仁组织区,是rDNA所在的位置。
二、核糖体
Robinson&Brown(1953)发现于植物细胞。
Palacle(1955)发现于动物细胞。
Roberts(1958)建议命名为核糖核蛋白(ribosome),简称核糖体。
核糖体是细胞内合成蛋白质的工厂,在一个旺盛生长的细菌中,大约有20000个核糖体,其蛋白占细胞总蛋白的10%,RNA占细胞总RNA的80%。
(一)、核糖体的结构
核糖体含40%的蛋白质、60%的RNA,蛋白按照一定的顺序与RNA结合,组成两个核糖体亚单体,其中RNAs是骨架结构,有些蛋白质不直接与RNA结合,而是结合在其它蛋白质组分上。核糖体中的蛋白质,rRNA以及其他一些辅助因子在一起提供了翻译过程所需的全部酶活性,这些酶活性只有在核糖体整体结构存在的情况下才具备。
不同来源的核糖体其形状,大小、化学组成稍有不同,通常根据沉降系数的不同分为70S和80S两种类型,70S核糖体存在于细菌,线粒体和叶绿体中,80S核糖体存在于真核生物的细胞质中。
所有的核糖体都是由大小两个亚基构成,核糖体的大小亚单位只有在以mRNA为模板合成蛋白质时才结合在一起,肽链合成终止后,大小亚单位又解离,游离于细胞质基质中。核糖体并不是单独工作的,而是由多个甚至几十个串连在一条mRNA分子上,称多聚核糖体(polyribosome),这样越长的mRNA可以结合更多的核糖体,提高了蛋白质合成的速度。
尽管原核生物与真核生物核糖体的蛋白质和rRNA差异很大,但结构总体相似,特别是负责与mRNA结合的小亚基。原核和真核细胞的rRNA都具有甲基化现象,这种甲基化与RNA的转录后加工过程的酶识别有关,另外原核5SrRNA和真核5.8SrRNA结构高度保守,常用于研究生物进化。
(二)、核糖体组装
编码rRNA的DNA片段称rRNA基因,它是重复的多拷贝基因,人的一个细胞中约有200个拷贝。rDNA没有组蛋白核心,是裸露的DNA节段,两个相邻基因之间为一段非转录的间隔DNA。
转录时,RNA聚合酶沿DNA分子排列,此酶由基因头端向末端移动,转录好的rRNA分子从聚合酶处伸出,愈近末端愈长,并且从左右两侧均可伸出,呈羽毛状(图12-29)。rRNA首先出现在纤维部,而后转向颗粒部。
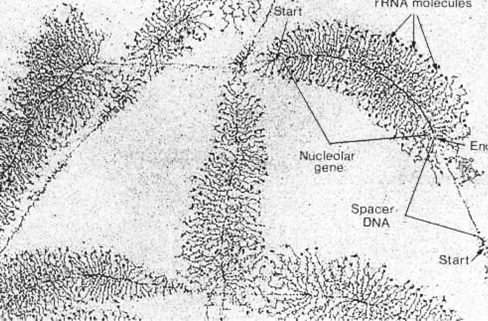
图12-29 rRNA的转录
纤维部的纤维状物质是新合成的45SrRNA,它与蛋白质形成RNP复合体,45SrRNA甲基化以后经RNA酶裂解为2个分子,18SrRNA和32SrRNA,后者再裂解为28SrRNA的5.8SrRNA。成熟的rRNA仅为45SrRNA的一半,丢失的大部分是非甲基化和GC含量较高的区域。
5SrRNA的基因并不定位在核仁上,通常定位在常染色体,5SrRNA合成后被转运至核仁区参与大亚基的装配(图12-30)。
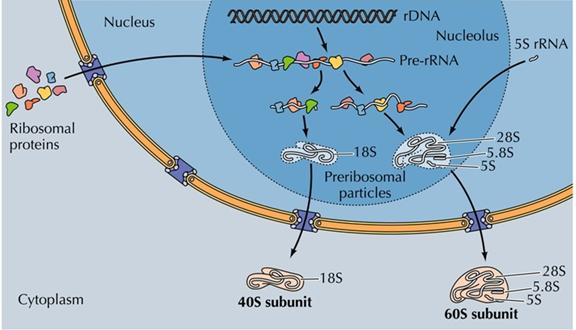
图12-30 核糖体的组装