|
��
������ �����뵰���ʸ�Ⱦ����
������Virus����һ���ϸ����̬�Ľ����������������ʽ֮������ʡ���������Ҫ������������С����ͨ���˾���������������õ羵���ܿ�������������һ�����͵ĺ��ᣬ��DNA��RNA��û�к����ֺ���IJ�������רӪϸ���ڼ�������������������ᵰ�ף�receptor binding protein����������ϸ������IJ����������ᣬ������Ⱦϸ����
һ����������̬�ͽṹ
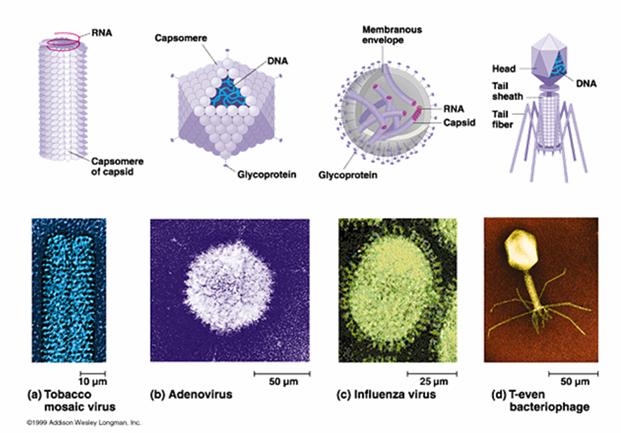
�����Ĵ�Сһ����10~30nm֮�䡣�ṹ���ɺ��ᣨDNA��RNA��о�͵������¿ǣ�capsid�������ɣ��ƺ��¿ǣ�nucleocapsid�����¿��б������������ø���������á����ֲ����������Ŵ���Ϣ����ͬ���ٵ�ֻ����3������Ŀɴ�300����ͬ�Ļ���
�����¿���һ�����ֵ�����ɣ���ɲ����¿ǵ��ǵ�λ�ƿ�����capsomer�����������γɲ���Ҫø�IJμӣ�ֻҪ�����߱�������͵����ʱ������װ�䣨self assembly���ɲ�������װ����ʽ�ж�ʮ����Գơ������Գƺ��϶Գ��������͡���ʮ����Գ��͵��¿ǵ����γɶ�ʮ���壬����������У������Գ��͵��¿ǵ������������������У����ύ֯�����У����϶Գ���Ϊͬʱ���л������ֶԳ�����ʽ�IJ�����ͼ3-21����
ͼ3-21
�����¿ǵ����з�ʽ
һ�������и�Ⱦ�ԵIJ���������������������Virion�����羵�۲���������̬�������Σ�Sphericity�������������Ͷ��ﲡ��Ϊ���Σ��缹������ײ�����ͼ3-21������������ٲ����ȣ���˿�Σ�Filament���������ֲ�ﲡ�������̲ݻ�Ҷ��������ͼ3-22�����������в�����ʱҲ��˿�Σ������Σ�Bullet-shape���������ӵ�ͷ�����Ȯ�����������Կ��ף�ͼ3-23�������ȣ�������Ϊֲ�ﲡ������ש�Σ�Brick-shape�������컨������ͼ3-24����ţ���粡���ȣ�������Σ�Tadpole-shape������һ��Բ�ε�ͷ��һ��ϸ����β��ɣ����ɾ�����ͼ3-25����������Ϊ��ʮ����Գƣ�������Ϊ�����Գƣ�������Ϊ���϶Գơ�
ͼ3-21 ��������ײ�����ͼƬ����http://www.denniskunkel.com/
ͼ3-22 �̲ݻ�Ҷ������ͼƬ����http://www.denniskunkel.com/
ͼ3-23 �����Կ��ף�vesicular stomatitis����������̬���Ȯ�������ƣ���Ϊ�ӵ��Ρ�ͼƬ����http://www.vetmed.ucdavis.edu
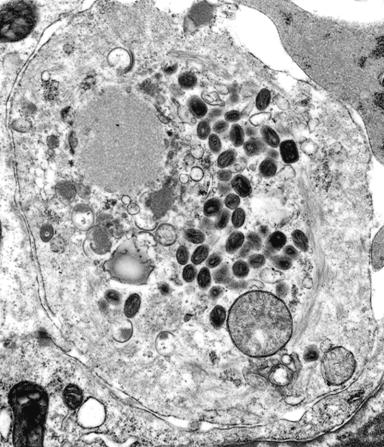
ͼ3-24�����컨������ͼ����Ⱦ�Ŀ�������ͼƬ����http://www.vetmed.ucdavis.edu
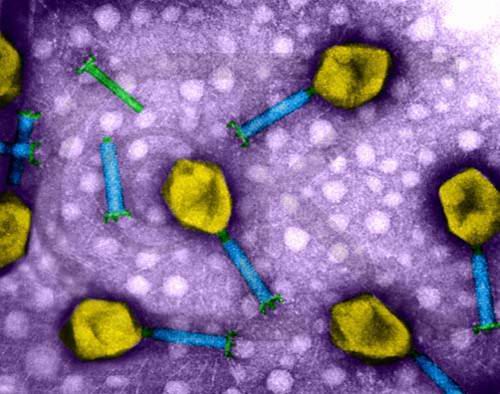
ͼ3-25 T4�ɾ��塣 ͼƬ����http://www.denniskunkel.com/
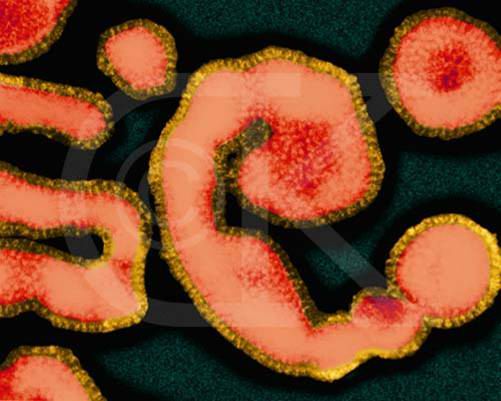
ͼ3-26 ˿״�б�Ĥ�IJ���������ð��������ͼƬ����http://www.denniskunkel.com/
�еIJ����¿���������һ�㱻Ĥ��viral envelope��ͼ3-26������㱻Ĥ�Dz�����������ϸ��ʱ�������ϵ�����ϸ������Ĥ����Ĥ�к��в����ںϵ��ף�viral fusion protein���������в����������ںϵ����ڲ�����������ϸ��ʱ���Źؼ����á�
����ֻ��������ϸ���Ժ�ű��ֳ��������������������ڿɷ�Ϊ�����Σ�һ����ϸ����Σ��Գ���IJ���������ʽ���ڣ���һ����ϸ���ڽΣ�����Ⱦ�Σ��ڴ˽��н��и��ƺͷ�ֳ����Ⱦ�ο�ʼʱ���������Ŵ��������¿����ͷų�����ע������ϸ���У�Ȼ���ڲ���������Ϣ��ָ�������£��γ��µIJ������ӡ�
���ݼ�����������ͬ�������ɷ�Ϊ���ﲡ����ֲ�ﲡ����ϸ�����������ɾ��壩�����ࡣ���ݲ��������ĺ�������ʺ�״̬��ͬ���ɽ�������Ϊ6�ࣺ
1)
˫����DNA��+mRNA�������ʣ����컨������T-ż���ɾ��塣
2)
����+DNA����DNA��+RNA�������ʡ�
3)
˫����RNA��+mRNA����������������²�����
4)
����+RNA����RNA��+RNA�������ʼ�������ײ�����
5)
����-RNA��+RNA��+�����ʣ������в����������в�������Ȯ������
6)
����+RNA��DNA����DNA��+mRNA�������ʣ�����ת¼������retrovirus���ֳ�RNA����������oncornavirus����
�����ಡ��
�ಡ���ڽṹ�ϱȲ�����Ҫ��û�е�������ǣ���Ϊһ��¶��RNA���ӡ��������Ǿ��и�Ⱦ���ã������ڲ������ʳ�Ϊ�ಡ����viroid�������Dz�����������Ⱦϸ����ֻ�е�ֲ��ϸ���ܵ����ˣ�ʧȥ��Ĥ���ϣ����Dz����ڹ���ֲ��������ֲ��䴫Ⱦ�����磬�����������ಡ������һ����359��������ĵ�����״RNA������ɣ�������һЩ�������С����ӳ�Լ40~50nm�����������¿ǵ��ס�
���������Ľ�����λ
�����Dz��������ಡ���������ж�����������ϳɵ����������Ƕ���ϸ���ļ��������ڽ����ϲ����ij��ֲ���������ϸ����������ǰ���ܿ�����������Ⱦɫ����������и��Ƶ�������plasmid������������DNA�͵ģ�Ҳ��RNA�͵ġ����벡������֮����Ҫ���ڣ�������רһ�ĺ�����������Ϊ���Ƶ���ʼ��λ�������ֲ�ͬ�ڲ������������쵰������ǣ�������һ����һ��ϸ�����ݵ���һ��ϸ������DNA���������Ϊ�¿ǵ����ʱ���Ļ���ʱ������ζ�Ų��������ˡ�
���������ּ䴫�ݺ������У�����������������������Ҫ���á����ڲ�������������ͬ����Ⱦɫ�����飬���Բ���������п���������һС������Ⱦɫ�壬һͬ���ݵ���һ��ϸ�����л����С��������ߣ�����DNA���ϵ�����Ⱦɫ���У�����������������һ���֣��ⲿ��DNA��Ϊǰ������provirus����ͨ������������ϸ��������䴫��DNA���еĹ��̳�ΪDNAת����DNA transduction���������﹤�̺ͷ�������ѧ�о��г�ͨ������;����ת��Ŀ�Ļ�������ijЩ������ϸ�����������Ҳ�����ձ����壬ͨ���Բ�������о������ڶ�������������⡣���磺
1.
����װ�䡣�����ɷ����ޣ��ṹ����ֻҪ�ɷ��뱸���������ˣ������Զ�װ����л��ԵIJ�����
2.
װ����Ϣ���Է��ӱ�����
3.
�Ŵ������ͳһ�ԡ�������ϸ��Я�����Ŵ���������ͬ�ģ�ʹ����ͬһ���Ŵ����롣
4.
���Ҹ��ơ���������ϸ�����ܹ����Ҹ��ƺͷ�ֳ����������������
�ġ������ʸ�Ⱦ����
1982��S��B��Prusiner�������Dz���Ϊʵ����ϣ���������������scrapie���IJ�ԭ����һ�ֵ����ʣ��������ᣬ����Ϊprion���⼴PROteinnaceous Infection ONly����Ϊ�����ʸ�Ⱦ���ӻ��ò�����Prusiner�����ָ�����ҽѧ��Ⱦ�ĸ����1997���ŵ����������ҽѧ����
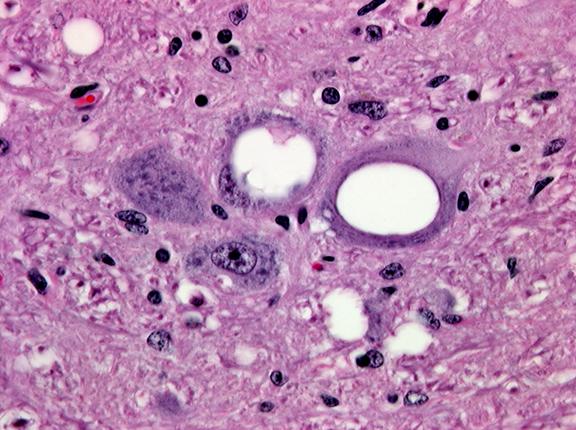
ͼ3-27��������֯�ĺ���״���� ͼƬ����http://www.vetmed.ucdavis.edu
����������������200�����ʷ����������ֲ��ͻ���������������ڼ�Ӳ�����ϴ�����壬�������������һ�ഫȾ�Ժ���״�Բ���transmissible spongiform encephalopathies��TSE������ţ������ţ����״�Բ���bovine spongiform encephalopathy��BSE��Ҳ���ڴ��༲����������1986�꣬������ţ��ι��������Ƿ���������϶�����Ⱦ����ţ���ڻ��ʼ���Ԫ���е��͵ĺ���״�˻���ͼ3-27��28�������ֵ�������amyloid�����׳�����ͼ3-28�����������������ơ�ͬ���͵�prionҲ��ʹ¹���������ӻ���������Ҳ�������Ƶļ�����
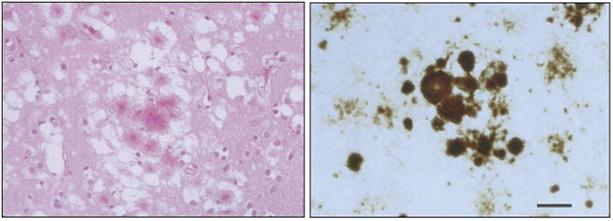
ͼ3-28
vCJD���˴�����֯��Ƭ������״���估��Χ�ij����ߣ��ҡ����������׳��������߱�ʾ50um������Stanley B. Prusiner 1997
Prion��һ�ֽṹ����ĵ����ʣ��Ը��º͵���ø�����н�ǿ�ĵֿ���������ת��ϸ���ڵĴ��������ĵ���PrPC��cellular prion protein����ʹPrPC�����ṹ���죬��Ϊ�����²����õ�PrPSc��scrapie-associated prion protein����
PrPC��������Ԫ������ϸ��������һЩϸ������������֬������ê�����ף�������Ĥ�ϵ�֬���У��Ե���ø�������У����ܺ�ϸ���ź�ת���йء�
PrPSc��PrPc��һ���ṹ������ͼ3-29��������253-4����������ɣ�������Լ33-37KD��������Prion������Ҷ�任���������������PrPc�ĸ��ṹ�о���43%�Ħ����������ٵĦ��۵���3%������PrPsc����34%��������43%�Ħ��۵������ﱻ��Ⱦ���������۵���PrPSc���ѻ�������֯�У��γɲ��ܵĵ��������׳�������������ø�ֽ⣬������ϸ��������Apoptosis����
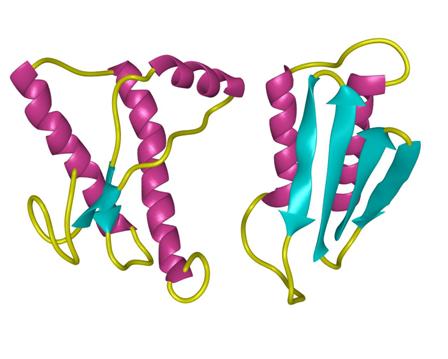
ͼ3-29 Prion�Ľṹģ�ͣ���PrPc����PrPSc ͼƬ����http://www.cmpharm.ucsf.edu/cohen/media/pages/gallery.html
����PrP���Ļ����ΪPrnp���û���λ���˵�20��Ⱦɫ��Ķ̱ۣ�С���2��Ⱦɫ�塣�ó�С���Prnp����С����������������������������ȫ���ߣ���������ܿ����ֹ���ʧ����С��Ƥ�����ϸ���˻���
Ŀǰ�Ե����ʸ�Ⱦ���ӵ���ֳ��ʽ�����ֽ�����ͼ3-30����һ�����۵�ģ�ͣ�refolding model������ΪPrPSc��������Ӱ��£�molecular
chaperone�������ã�����PrPc�������ϣ���ʹPrPcת���PrPSc���Ӷ��γ���PrPSc�����壬����һ��PrPSc���Ӿͱ����2��PrPSc���ӣ���˱������ѡ���һ�ֽ����Ǿ���ģ�ͣ�Seeding model������ΪPrPc���ӱ�������PrPScת�������һ��ƽ�ⷴӦ����PrPSc������һ�����ȶ�PrPc�Ĺ����γɵ��������׳�����Ȼ�����Ѻ��ֱ���µľ��֡�
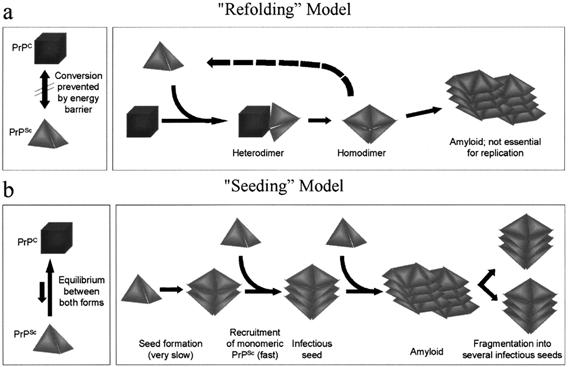
ͼ3-30 PrPSc����ֳ ͼƬ����Frank
L. Heppner��2001
�����ʸ�Ⱦ���ӵ���ֳ�Ȳ������ڻ�����ֱ��Ҳ�������������ӣ����������������ӵĹ�����ת����ɵģ���������ò�����Ŀǰ��֪������PRION������Ҫ�У�
1.
��-�Ŷ��ϲ���Creutzfeldt�CJakob
disease��CJD����Cruetzfeldt��Jakob 1920�귢�����������ߣ�������60�����ϵ��ˣ�������PrP��������������ġ�
2.
�����Ϳ�-���ϲ���vCJD�������߶���������CJDδ�����ֵ�����Σ�Ϊʮ��������ʮ��������ˣ�������ȡʳ��ţ��Ʒ����Ⱦ���������ȳ�������֢�IJ�״���̶��������ߣ������־����ϰ��ȳմ�֢״�����������
3.
GSS�ۺ�����Gerstmann-Straussler Scheinker disease��������һ���Ŵ��ĵ������Բ�����Prnp����ȱ������PrP����102λ�����ᱻ������ȡ����117λ���Ӱ��ᱻ������ȡ����
4.
��³����Kuru�����������¼�����һ����Fore�IJ��䣬�����˳���kuru���⼴���������˴�����Ǹ�Ů��С������֢�����ﺬ��������ʶ�ؿ�Ц�����ʡ���²�������һ������ҽ��D. C. Gajdusek���˵��أ���������ĸ�Ů��С�����г�����ʬ���ϰ�ߣ�����ܵ���Ⱦ��
5.
�����Լ�����ʧ��֢��Fatal familial
insomnia��FFI����Ҳ��һ���Ŵ��Լ�����Prnp������죬PrP����178λ���춬�������춬����ȡ�������ߵ���Ҫ֢״��ʧ�ߣ�����CJD��֢״��
���ڵ����ʸ�Ⱦ��������ļ�����Ŀǰ��û����Ч�����ƴ�ʩ�����൰���к�ǿ�ĵֿ������Կ����غ������������У�134-138������1h�IJ�ţ����֯�Ƚ����Լ�10%�������̶ֹ����IJ�������֯�����и�Ⱦ�ԡ�
�ݱ�������1996������������106�˵��˷�ţ�������н������˻����š�
�� |